
尺寸排阻色谱法 (SEC) 是用来测量蛋白质分子量(MW 或摩尔质量)的一个惯用方法。 传统上,该方法涉及对比样品的色谱柱保留体积与一系列已知分子量的标准品的色谱柱保留体积。 重要的是,SEC 的分离过程是由分子的流体力学体积控制的,而不是分子量。因此,该方法产生的结果仅仅是相对于用于校正系统的标准品而言的。 样品的“相对”分子量与实际分子量的相近程度,是由标准品与分析物结构的相似度决定的。 这表明蛋白质样品结构与蛋白质标准品结构的偏差会对结果的准确性造成负面影响。 此外,某些蛋白质会与色谱柱基质发生相互作用或在洗脱期间改变其流体力学体积。 在这些情况下,无法依赖保留体积来计算所测量分子的分子量。
色谱系统新增的先进检测器可在不依赖任何标准品或样品洗脱体积的基础上测量样品的绝对分子量。
在此应用说明中,我们将看到两个实例,其样品的保留体积不可用于获得样品分子量的准确测量。 我们还将说明使用这些先进的检测器可获得的其他信息。
此应用说明所介绍的两个应用中,所用的仪器为 Viscotek TDAmax 系统,其中包括一个连接至 GPCmax 脱气装置、泵和自动进样模块的三重检测器阵列 (TDA),并配备结果部分中指定的 SEC 色谱柱。 牛血清白蛋白 (BSA) 进样用于确定检测器间延迟体积、校正常数和谱带扩展校正因子。
OmniSEC 软件用于控制整个系统并采集来自 TDA 系统的 5 个检测器的信号:紫外可见光分光光度计、测量静态光散射强度的 7° 小角度光散射 (LALS) 和 90° 直角光散射 (RALS) 检测器、示差折光 (RI) 检测器和惠斯登电桥压差 (DP) 粘度计。 OmniSEC 用于根据上述测量值(RI、UV、LALS、RALS 和 DP)计算蛋白质的绝对分子量 (MW) 和特性粘度 (IV)。
理想状态下,使用 SEC 分析样品时不应有样品与色谱柱基质的相互作用。 然而,在实际实验中这并非总是可以避免的。 使用先进的检测器系统的优点是,即使出现了样品与填料间的相互作用,仍然可获得绝对分子量值。
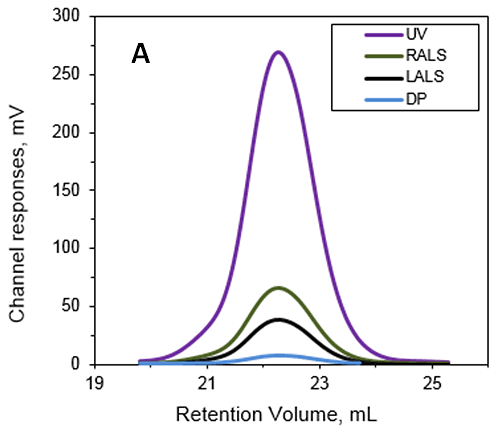
本实验研究了 Anthrolysin (ALO),这是一种由炭疽杆菌分泌的成孔胆固醇依赖型细胞裂解素 (CDC)。 研究表明,ALO 在炭疽热的发病机制中产生了一定的作用。 研究中使用 Superdex 200 (GE Healthcare) 与含 20mM 三羟甲基氨基甲烷、150mM 氯化钠的缓冲液(pH 值为 7.3)进行了 SEC 实验。 Viscotek TDA 用于测定溶液中 ALO 的分子量和特性粘度,结果如图 1 所示。
|
|
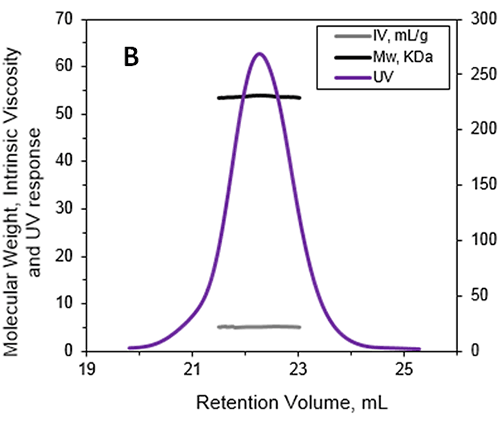
计算得出的绝对分子量为 53.6 kDa,表明 ALO 在溶液中以单体存在。 所记录峰的保留体积为 22.2 mL,这可能是由于样品与填料之间的相互作用导致的。 如果使用的是传统色谱校正方法,这将相当于仅为 15-20 kDa 的分子量。 这说明使用光散射检测器来防止低估样品分子量是很有必要的。 关键在于,并不是每次都可以确定样品与填料之间是否发生了相互作用。 这意味着,若是本次研究中未使用这些先进的检测器,对蛋白质分子量的低估多半会被忽视。
除了测量 ALO 的绝对分子量以外,还使用了一个在线粘度计测量了 IV 值。 IV 用于衡量溶质中的单分子对溶液整体粘度的贡献。 IV 与物质的分子密度成反比,若蛋白质结构、形状和水合作用(即灵活性)发生了变化,就会导致蛋白质体积的改变,并最终影响密度变化。 ALO (53.6 kDa) 的 IV 值测量结果为 5.1 mL/g。 作为对照,BSA 单体 (66.5 kDa) 的 IV 值约为 4 mL/g。 通过将 IV 和流体力学半径数据相结合,可预估 ALO 的形状和水合作用(见参考文献 1 和 3):ALO 的水合作用较弱,表明该蛋白质是折叠的;ALO 的形状是细长的,这与 X 射线结构的锥形形状吻合。
传统 SEC 实验的一个最大的局限性在于无法说明标准品和感兴趣样品之间的结构差异。 本例主要针对 Bcl-2 蛋白家族。 这些蛋白质是对内部和外部凋亡信号非常敏感的分子传感器,在调节细胞凋亡方面发挥着重要作用。 由于与多种疾病有关的蛋白质错误折叠,其聚集会导致淀粉样纤维的行成。 淀粉样沉积的形成已被证实与神经退行性疾病的异常细胞凋亡有关。 在体内,细胞凋亡控制依赖于 Bcl-2 家族成员间的相互作用以及异聚体的形成。
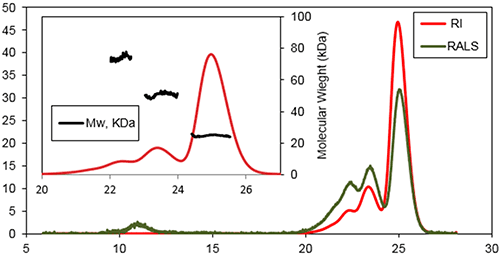
本研究重点关注 Bcl-2 蛋白(通过在 37°C 下进行蛋白培养生成)在体外的聚合物形成。 SEC 用于表征在培养一天和一周后的聚集过程中出现的早期事件。 图 2 是在 37°C 下培养一天后的典型三重检测色谱图。 研究中使用 Superose 6HR (GE Healthcare) 与含 20 mM 磷酸钠及 150 mM 氯化钠的缓冲液(pH 值为 8)进行了 SEC 实验。
|
检测到的三个主要品种保留体积如下:22.6、23.6 和 25.1 mL,分子量分别为 74±3 kDa、51±2 kDa 及 25±2 kDa。 研究发现,该样品的主要种群为蛋白质单体状态的较新品种。 在三聚物的峰上可检测到一个肩峰值,很可能是四聚物的一个小种群。 在洗脱至 11 mL 时,检测到另一个品种。 该品种是由光散射检测器所记录的,而非示差折光检测器。 这是有着极高分子量的蛋白聚合物的典型反应,所以反射了大量的光,而浓度却较低。
如果该研究使用的是传统色谱校正方法,Bcl-2 单体 (25.1 mL) 的表观分子量将仅为 40 kDa。 该数据表明,尽管 Bcl-2 是单体结构,其流体力学半径 (RH) 却比预期的要高。 假设样品与色谱柱基质之间没有产生相互作用,由于高蛋白质水合作用和/或球形偏差,就有可能会出现这种效应。 Bcl-2 是一个球状蛋白质,有八个 α 螺旋和一个将第一个 α 螺旋连接至蛋白质核心的弹性环。 弹性环在样品洗脱图中发挥了极其重要的作用,使得该分子量的蛋白质流体力学半径比小型蛋白质预期的要更高。
研究对在 37 °C 下培养一周后的样品也进行了分析。 在这段延长的培养期内,研究发现有更多的聚合物形成。 SEC 实验表明,纤维生长的第一步是形成小的聚合物,多半是二聚物和三聚物,然后再组合形成更大的聚合物。
SEC 是研究蛋白质及其功能的一个至关重要的工具。 在此我们讨论了在测量样品分子量时依赖传统色谱校正方法的缺点。 使用多检测系统表明,我们可以计算蛋白质及其寡聚物的绝对分子量,无需依赖保留体积或任何用于分子质量校正的标准品。 这一先进的方法也已被证实可用于辨别样品内聚合物状态、聚合程度、水合作用或形状之间的差异。
Bourdeau, R. W., Malito, E., Chenal, A., Bishop, B. L., Musch, M. W., Villereal, M. L., Chang, E. B., Mosser, E. M., Rest, R. F., and Tang, W. J. (2009) Cellular functions and X-ray structure of anthrolysin O, a cholesterol-dependent cytolysin secreted by Bacillus anthracis. J Biol Chem 284, 14645-14656.
Chenal, A., Vendrely, C., Vitrac, H., Karst, J. C., Gonneaud, A., Blanchet, C. E., Pichard, S., Garcia, E., Salin, B., Catty, P., Gillet, D., Hussy, N., Marquette, C., Almunia, C., and Forge, V. (2012) Amyloid fibrils formed by the programmed cell death regulator Bcl-xL. J Mol Biol 415, 584-599.
Karst JC, Sotomayor-Pérez AC, Ladant D, Chenal A. (2012) Estimation of intrinsically disordered protein shape and time-averaged apparent hydration in native conditions by a combination of hydrodynamic methods. Methods Mol Biol. 2012;896:163-77.