Arginine has been used widely over the last 20 years in protein refolding experiments and as a biopharmaceutical excipient. Whilst being an effective aggregation suppressor, the additive has been shown to have little effect on the conformational stability of proteins [1]. Despite a great deal of research, the mechanism of aggregation-inhibition by arginine is still not fully understood. It has been suggested that arginine binds to a protein's surface and increases intermolecular repulsion, as the guanidinium group on arginine's side-chain is positively charged at neutral pH.
Much of the research carried out into the influence of arginine on protein stability has involved the use of spectroscopic techniques that analyze the bulk properties of the sample. On the other hand, light scattering techniques such as Dynamic Light Scattering (DLS) and Static Light Scattering (SLS) are capable of detecting the formation of trace amounts of oligomer, allowing elucidation of the earliest stages of the aggregation process.
This application note describes the characterization of a model protein (lysozyme) in the presence and absence of arginine using the array of techniques provided by the Zetasizer Nano ZSP. SLS is used to assess the temperature at which lysozyme aggregation begins, with temperature control provided by the instrument's peltier elements. DLS is used to assess the various particle sizes present in the two formulations at different temperatures, whilst measurements of zeta potential are used to assess the effect of arginine on the charge state of the protein.
|
During this study, samples were heated from 20°C to 90°C, with measurements acquired at temperature intervals of 0.5°C. A single DLS measurement consisting of 15 runs, each lasting 10 seconds, was performed at each temperature, following a 30 second equilibration period. Samples were held at each temperature, therefore, for 3 minutes in total. These thermal profile experiments were carried out in triplicate for each of the two formulations, allowing the repeatability of both the DLS measurements and the temperature ramp to be assessed.
Electrophoretic mobility at 25°C was measured with a disposable folded capillary cell using the diffusion barrier method [2,3] in order to minimize the potentially denaturing effect of contact between the sample and the cell electrodes. The Zetasizer was set to automatically select the optimum voltage and attenuation level for the electrophoretic mobility measurements.
One sample consisted of 2 mg/mL lysozyme in a 10 mM phosphate buffer at pH 7, whilst the other consisted of 2 mg/mL lysozyme in a 10 mM phosphate buffer containing 50 mM arginine at pH 7. Both samples were filtered using a pore size of 20 nm immediately after preparation.
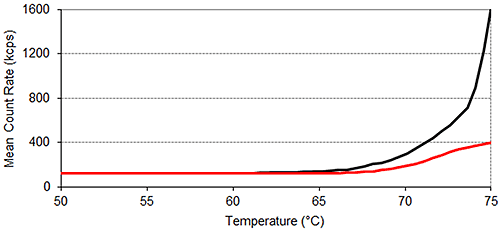
Figure 2 shows the results of SLS monitored thermal profile experiments. Figure 2 clearly demonstrates that in the absence of arginine, lysozyme starts to aggregate at a lower temperature than it does in the presence of arginine. The mean count rate between 25°C and 50°C averages 121 kilo counts per second (kcps) for lysozyme in the presence of arginine, and 123 kcps for lysozyme in the absence of arginine. This value rises above 250 kcps at 71.5°C ± 0.0°C in the presence of arginine and at
69.5°C ± 0.0°C in the absence of the additive. Above 70°C, the presence of arginine severely inhibits the rise in light scattering intensity, a measure of the average particle size of a sample, with temperature. The data presented in Figure 2 therefore suggests that arginine suppresses the aggregation of lysozyme.
|
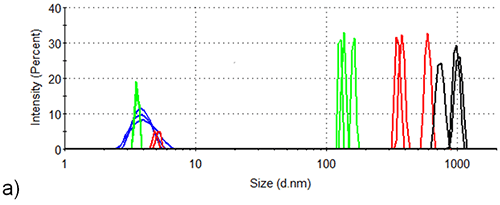
Figure 3 shows the intensity-based size distributions in the two samples generated by DLS measurements at three different temperatures during the thermal profile runs shown in Figure 2. At 25°C, the intensity size distributions of both samples reveal the presence of monomeric lysozyme with a hydrodynamic diameter of 4 nm, no aggregates being detected. As the temperature rises to 71.0°C, the intensity size distributions of lysozyme in the absence of arginine become dominated by aggregates with hydrodynamic diameters of 100 nm - 200 nm, while those size distributions for lysozyme in the presence of arginine are contributed to equally by lysozyme aggregates and monomers. This is consistent with arginine being able to delay the onset of aggregation in protein formulations.
An extra 5°C rise in temperature, to 76.5°C, leads to the development of larger aggregates (300 nm - 700 nm hydrodynamic diameter) in the absence of arginine, while in the presence of arginine the largest particles detected are < 300 nm hydrodynamic diameter. In addition, a lysozyme aggregation peak with a hydrodynamic diameter of ~ 20 nm is also detected at 76.5 °C in the formulation containing arginine. An aggregate of this size is not detected at any point during the aggregation process of lysozyme in the absence of arginine.
As the temperature rises to 84.0°C, aggregates in the formulation which does not contain arginine continue to grow, particles with a hydrodynamic diameter > 600 nm being the only species in the intensity analysis at this point. In the presence of arginine, however, the maximum aggregate size is < 500 nm, the aggregates showing very little growth in hydrodynamic diameter between 71°C and 84°C. The contribution of the 20 nm peak (first detected at 76.5 °C) to the intensity size distribution, relative to that of the larger peak, decreases as the temperature rises from 76.5 to 84.0 °C, suggesting aggregation of the 20 nm diameter oligomers over this range. Clearly, the presence of arginine in formulation buffers influences the mechanism of aggregation.
|
|
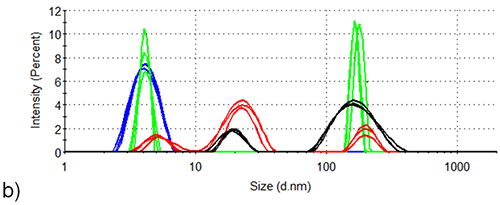
Figure 4 shows the volume size distributions of lysozyme in the presence and absence of arginine at 84.0°C, the distributions calculated from the corresponding intensity distributions (Figure 3). Table 1 displays the statistical description of these volume size distributions. In the absence of arginine, all particles making a measurable contribution to the protein-occupied volume of the sample have hydrodynamic diameters > 600 nm. In the presence of arginine, particles of 10-30 nm account for the vast majority of the protein-occupied volume, with larger particles of 90 nm - 200 nm diameter occupying a volume < 1/40 of that occupied by the smaller aggregates.
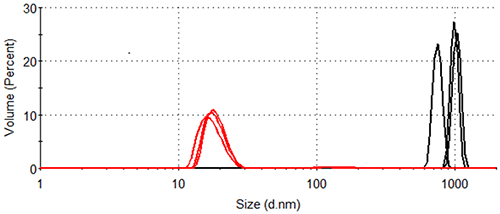
|
| Formulation | Hydrodynamic Diameter (nm) | % Contribution to Volume Distribution |
|---|---|---|
| Lysozyme in pH 7.0 10 mM phosphate buffer | 924 ± 153 | 100 |
| Lysozyme in pH 7.0 10 mM phosphate buffer containing 50 mM arginine | 18.1 ± 0.8 | 97.9 ± 0.4 |
| 161 ± 6 | 2.1 ± 0.4 |
In order to test the hypothesis that the effect of arginine on protein aggregation is due to binding of the amino acid to the protein, leading to increased intermolecular repulsion, the zeta potential of native lysozyme was measured both in the presence and in the absence of arginine. The data from electrophoretic light scattering (ELS) measurements are shown in Table 2. Arginine lowers the zeta potential of native lysozyme at 25°C. Such charge-screening by arginine would be expected to decrease intermolecular repulsion, rather than increase it. Table 2 suggests, therefore, that the inhibition of aggregation by arginine is not due to its effect on the native protein. This finding would support the alternative hypothesis that arginine binds to the hydrophobic areas of protein which become exposed upon denaturation, acting as a charged protectant of these aggregation hotspots [4,5].
| Formulation | Zeta Potential (mV) |
|---|---|
| Lysozyme in phosphate buffer | 7.0 ± 0.1 |
| Lysozyme in phosphate buffer containing 50 mM arginine | 4.6 ± 0.5 |
Orthogonal analysis of protein aggregation can be carried out quickly and simply using the Zetasizer Nano ZSP. The sensitivity of the instrument to trace amounts of aggregate allows the early stages of aggregation to be characterized. SLS was used to demonstrate that the presence of arginine increases the onset temperature of lysozyme aggregation by 2°C, confirming that the additive is able to inhibit the earliest stages of aggregation.
DLS was used to show that arginine not only delays the onset of aggregation, but also influences the pathway via which aggregation proceeds. Lysozyme in the absence of arginine simply forms aggregates that increase in hydrodynamic diameter as the temperature increases, particles of almost 1 µm being formed by the time the temperature has risen to 84.0°C. In the presence of arginine, however, aggregates remain < 500 nm throughout the thermal profile, with a slower lysozyme aggregation process that leads to the formation of aggregates with hydrodynamic diameters of 10 nm - 30 nm also detected. Volume distribution analysis was used to show that, in the presence of arginine, the smaller aggregates accounted for a much larger proportion of the total protein volume than the larger aggregates, even at 84.0°C. Such information is highly relevant to developers of biopharmaceuticals (arginine is often included in bioformulations), given the effect that aggregate size and concentration may have on the immunogenic potential of protein-based drug products [6].
Finally, ELS was used to demonstrate that the aggregation-inhibiting effect of arginine is not due to any effect of the additive on the net charge of the protein in its native state. This supports the theory that the arginine inhibits aggregation by increasing the intermolecular repulsion of the denatured protein, rather than that of the native protein.
